
| |
| ABOUT APRESLABS |
| INSECTICIDE RESISTANCE |
| SERVICES |
| APRESLABS BLOG |
| CONTACT |
| TARGET-SITE RESISTANCE | |||||||||
|
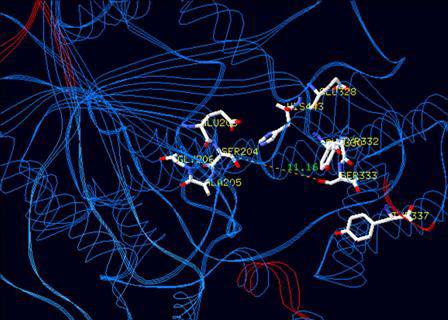
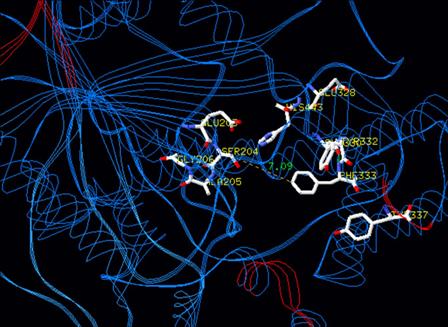
PUBLICATIONS PATENTS TEMPORAL SYNERGISM |
||
|
ApresLabs Ltd | |||||||||||||||||||||||||||||||||||||
| SPECIALISTS IN PESTICIDE RESISTANCE | ||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||
ApresLabs Ltd | tel: +44 (0)1582 763133 ext 2483 | Email: Graham.Moores@apreslabs.co.uk |